







































An Observed Reduction In Wine Cineole Content
August 26, 2024
Terpenes are common chemicals found naturally in wine and they occur in many forms, some examples of which are depicted in Figure 1. In general they exhibit pleasant floral and fruity odours. For a broad overview of these important wine aromatic contributors see Waterhouse et al, 2016.
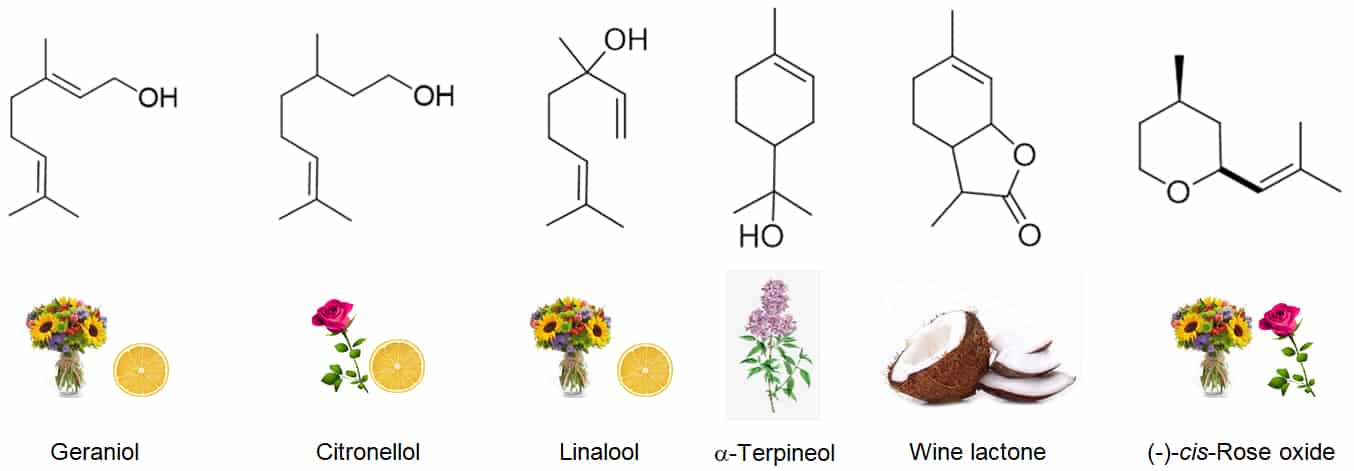
Figure 1: Examples of some of the terpenes found in wine, depicting chemical structures and associated aromatic descriptors.
Not all terpenes have obviously pleasant odours, however, with some bordering on medicinal. Perhaps the best example of this are the terpenes a- and b-pinene, which are the main aromatic constituents of turpentine, a solvent made from coniferous trees that is found in most garages.
Another, more pertinent example is the eucalyptus-smelling terpene sometimes observed in Australian wines (and those overseas if eucalypt trees are grown near vineyards (Capone et al, 2012) which goes by a few names. It is used somewhat in the cosmetics industry and its presence in the air throughout the Blue Mountains west of Sydney provided the origin of their name. In chemical nomenclature it has an ugly name that is often reduced to the trivial name of 1,8-Cineole (or even simply “cineole”) for convenience, as well as eucalyptol. It is a monoterpene cyclic ether found principally in eucalypt leaves, but also in other plant species such as Salvias and some herbs. The molecule’s internal plane of symmetry renders it achiral (Figure 2), meaning there is only one structural form (i.e., no enantiomeric left- and right-handed forms), although the structural isomer 1,4-Cineole also exists but is typically found to much lesser extents. It is usually found in red wines rather than white, presumably due to the longer skin contact and maceration associated with red wine production.
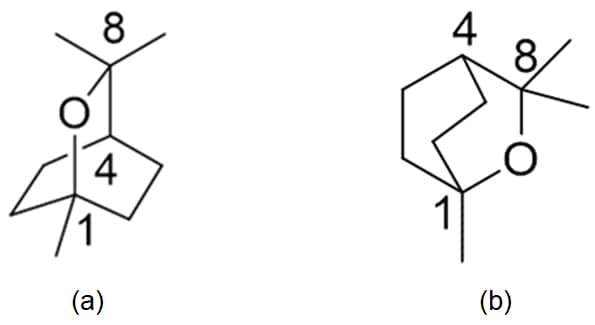
Figure 2: 1,8-Cineole structure, depicted from different perspectives, with (a) illustrating the molecular internal plane of symmetry (along the 1-4-8 axis) and (b) showing a bird’s-eye view above the heterocyclic ring. Selected carbon numerical assignments are indicated.
Terpenes are common in nature and, as a result of their low molecular polarity (Bowyer, 2003), are typically very volatile and are thus pungent in wine, often with low detection thresholds. Waterhouse et al (2016) report the cineole threshold as 1.1 ug/L with a range of 0-33 ug/L. The presence of any terpene in wine is of interest to most winemakers, but this is especially so for cineole for three primary reasons. Firstly, it is a non-varietal character (Bowyer, 2018), not being intrinsically found in grapes or wine unless eucalypt trees are growing nearby.
Secondly, cineole is very potent (viz. detection at 1.1 ug/L), so even a small amount can have a strong impact on the aromatic profile of a wine. Thirdly, some people do not like the aromatic contribution of cineole to wine. The latter is a personal choice of course, and is thus open to argument. Some winemakers advocate the character, but cineole is certainly a non-varietal character, as it is not found in wines made from grapes grown in the absence of eucalypt trees. There has also been at least one instance of successful litigation between a US wine producer and the owner of an adjoining property that had eucalypt trees planted on its boundary, where cineole from the trees was found to be undesirably present in the producer’s wine. This constitutes fairly clear evidence that the winemaker in question did not like the cineole character in his wine!
Cineole is, however, arguably a regional character (albeit non-varietal) in parts of Australia, notably Coonawarra and Langhorne Creek, amongst others, presumably due to the prevalence of eucalypt trees in proximity to the vineyards in these areas. Ultimately, one question often arises in discussions about this character in wine: can it be lowered? This article seeks to present an observation that may be of interest to those posing this question.
Trial background
The kinetic results of a yeast/bacteria coinoculation trial conducted in 2020 have been published previously (Bowyer & Mennesson, 2021). he focus of that trial was the investigation of early coinoculation of bacteria as a means of improving primary and secondary fermentation kinetics relative to late coinoculation (i.e. bacterial addition at the press tray). There are pros and cons to both approaches, but early coinoculation is gaining increasing favour with Australian winemakers.
A kinetic comparison of the two fermentations in the 2020 trial is reproduced in Figure 3. As this was a winery trial no replicates were run, however the trial was repeated in 2021 under the same conditions using the same fruit source, consumables and methodologies. The sluggish completion of primary fermentation under yeast strain B (solid blue trace, Figure 3) was the impetus for this trial. Further results pertaining to that trial are given in the original publication (Bowyer & Mennesson, 2021).
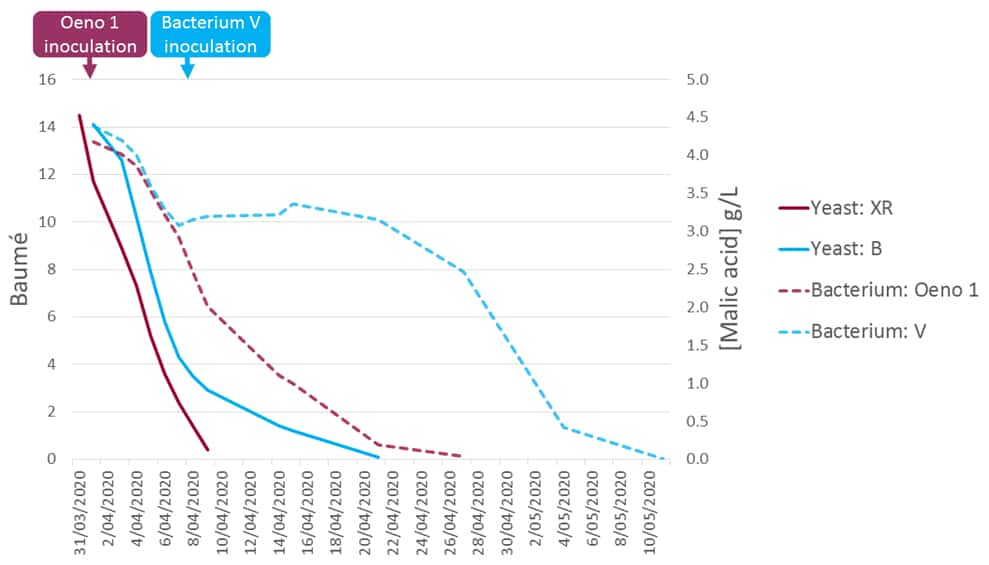
Figure 3: The kinetics of parallel fermentations conducted in 2020 using different winemaking protocols on common fruit. Purple traces indicate Excellence XR yeast (solid) and the Oeno 1 bacterium (dashed) under early coinoculation, and blue traces indicate yeast Strain B (solid) and bacterium V (dashed) under late coinoculation. Bacterial inoculation timing is indicated on the chart, for Oeno 1 (early) and Strain V (late).
When the two wines were compared in a tasting in June, and later again in December 2020 some significant organoleptic differences were noted. Firstly, the early coinoculation wine (yeast XR/bacterium Oeno 1) was more fruit-forward, which is typical for this technique. Secondly, the late coinoculation wine (yeast B/bacterium V) exhibited some volatile acidity, which is perhaps not unexpected given the difficulty of B completing primary fermentation (Figure 3, solid blue trace).
In addition to these, another difference was observed: the cineole levels of both wines. When this was articulated, both winemakers involved in the trial indicated that they too had observed this difference. This apparent difference was filed under “interesting”. The 1,8-Cineole levels of the 2020 wines were quantified by the AWRI in early 2021. As noted previously, this trial was repeated in 2021 using the same fruit and again a perceived difference in cineole levels was observed in the final wines, whereby the early coinoculation XR/Oeno 1 wine exhibited lower cineole on the nose, pushing this serendipitous outcome of the trial from the realms of “interesting” to “must investigate further”. Again, making use of the AWRI analytical service, the 2021 samples were submitted for analysis in September 2021.
Cineole quantification
Figure 4 indicates the levels of 1,8-Cineole measured in the respective wines for each vintage.
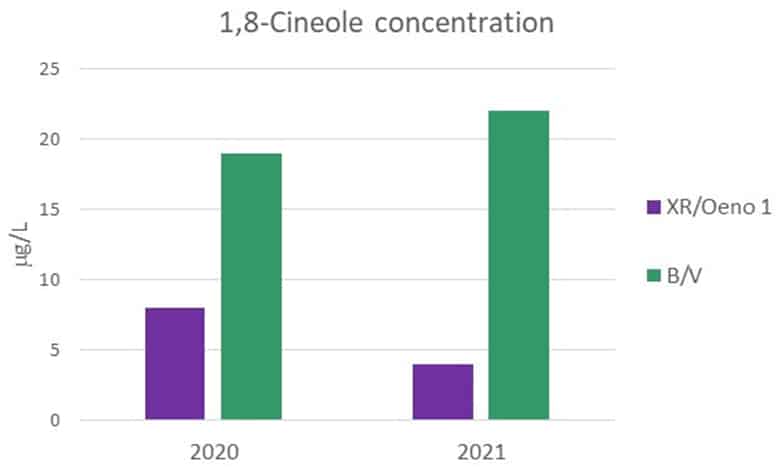
Figure 4: 1,8-Cineole concentrations in the trial wines from the 2020 and 2021 vintages, reported in ug/L.
Plausible cineole variance causatives
In terms of trial parameters, the variables are few. It should be remembered that this was simply a winery protocol comparison, and not a fully controlled experiment. Fruit and general winemaking parameters were uniform between the parallel fermentations. Different yeast (XR and Strain B) and bacteria (Oeno 1 and Strain V) were used in the fermentations, and different inoculation timing was employed (i.e., early vs. late coinoculation respectively). Oenostim rehydration nutrient was used only on the XR yeast in 2020, but in 2021 it was used for both fermentations, so that can be eliminated as a possible source of the cineole concentration variance.
If the timing of the coinoculation (i.e., early vs. late) was the cause of the cineole variance in the wines it would be reasonable to expect the ratio of cineole in each wine to have remained the same between 2020 and 2021, if not the absolute concentrations (which may vary according to the season), but this was not observed. In the 2020 wines the cineole ratio was 1 : 2.4 (XR/Oeno 1 : Strain B), but in the 2021 wines this ratio increased to 1 : 5.5, suggesting that bacterial inoculation timing is not likely to be causative.
Currently, the best working theory is that one (or both) of the microorganisms in the yeast/bacterium couple was responsible for the cineole variance. Given that the cineole molecule has very low molecular polarity it is possible, to some extent at least, that it is adsorbed into the cell membranes of both or either the XR yeast or Oeno 1 bacterium, and once racked the lees would thus remove some of the cineole. This adsorption affinity could be expected to vary between organism strains, and this could explain the observed variance. The yeast is the more likely adsorption candidate in this case due to the higher thickness of the yeast cell wall versus the bacterial cell wall (Grbin, 2023), but this and other parameters would need to be evaluated rigorously to confirm the working theory. It is also possible that the cineole variation was due to partial metabolism, although this seems less likely. Discussion has been initiated in this regard.
Summary
Many winemakers do not like the character imparted by eucalyptus trees to their wines and cineole, the chemical responsible for this aroma, is not easy to remove once present in a wine. A serendipitous observation in an unrelated trial in 2020 appears to have yielded an approach that can potentially decrease the amount of cineole in wine. Given that cineole is a non-varietal character, this can lead to wines of higher varietal typicity. Research is required to validate the theory behind the possible mode of action in this case and fully quantify the effects.
By Dr Paul Bowyer
References
Bowyer, P. K. and Mennesson, A. (2021) Coinoculation of yeast and bacteria, The Australian and New Zealand Grapegrower and Winemaker, January Issue, 684, 46-53.
Bowyer, P. K., Chancholle, L., and Mennesson, A. (2018) Understanding yeast aromatic expression: is my wine varietal?, The Australian and New Zealand Grapegrower and Winemaker, April Issue, 651, 54-56.
Bowyer, P. K. (2003) Molecular polarity – it’s behind more than you think, The Australian and New Zealand Grapegrower and Winemaker, November issue, 89-91.
Dimitra L. Capone, David W. Jeffery, and Mark A. Sefton (2012) Journal of Agricultural and Food Chemistry, 60 (9), 2281-2287
DOI: 10.1021/jf204499h
Grbin, Paul (2023), personal communication.
Waterhouse, A.L., Sacks, G.L. and Jeffery, D.W., 2016. Understanding wine chemistry. John Wiley & Sons, 68-78.
BHF wishes to thank The Australian and New Zealand Grapegrower and Winemaker for permission to reproduce this article. Subscription information can be found here.